ARTIGOS ORIGINAIS
Branqueamento de corais e outros cnidários bentônicos no costão rochoso da Praia do Forno (Arraial do Cabo, RJ) durante as anomalias térmicas das águas superficiais do oceano ocorridas nos meses de fevereiro e maio de 2019
Coral and other benthic cnidarians bleaching on the rocky shore of Forno Beach (Arraial do Cabo, Brazil) during the thermal anomalies of the ocean water surface in February and May 2019
Blanqueamiento de corales y otros cnidarios bentónicos en la orilla rocosa de la Praia do Forno (Arraial do Cabo, Brasil) durante las anomalías térmicas de las aguas superficiales del océano ocurridas en los meses de febrero y mayo de 2019

Branqueamento de corais e outros cnidários bentônicos no costão rochoso da Praia do Forno (Arraial do Cabo, RJ) durante as anomalias térmicas das águas superficiais do oceano ocorridas nos meses de fevereiro e maio de 2019
Vértices (Campos dos Goitacazes), vol. 23, núm. 2, 2021
Instituto Federal de Educação, Ciência e Tecnologia Fluminense

Esta obra está bajo una Licencia Creative Commons Atribución 4.0 Internacional.
Recepción: 03 Octubre 2020
Aprobación: 06 Abril 2021
Resumo: O presente estudo objetiva descrever as ocorrências de branqueamentos dos corais e de outros cnidários bentônicos no costão rochoso da Praia do Forno (Arraial do Cabo, RJ), observadas em fevereiro (verão) e maio (outono) de 2019, relacionando-as com possíveis eventos de aquecimento das águas superficiais do oceano. As cartas de anomalias térmicas foram obtidas pelo INMET e os trabalhos de campo foram desenvolvidos baseados no Protocolo AGRRA. Foram utilizados dois transectos de 10 m de comprimento, colocados de forma contínua e paralelos ao eixo maior do costão rochoso, onde foram distribuídos quadrats de 36X36 cm, nas posições demarcadas em: 1, 3, 5, 7 e 9 m ao longo destes transectos e fotografados para obtenção dos dados. Foram calculados: o número total de colônias, número de colônias branqueadas e a média da área de cada espécie de cnidário bentônico afetada por diferentes intensidades de branqueamento (fraco ou forte). Quatro espécies de cnidários bentônicos foram identificadas, dentre estas, Millepora alcicornis e Palythoa caribaeorum sofreram branqueamento forte e fraco, com aumento principalmente do branqueamento forte no mês de maio – período antecedido, por anomalia térmica de valor 2,0 ºC durante todo o mês de março e na segunda quinzena de abril; enquanto Siderastrea stellata . Zoanthus sp. não sofreram branqueamento.
Palavras-chave: Branqueamento de coral, Anomalia térmica, Praia do Forno, Arraial do Cabo, Costão rochoso.
Abstract: The present study aims to describe the occurrences of coral bleaching and other benthic cnidarians on the rocky shore of Forno Beach (Arraial do Cabo, RJ), observed in February (summer) and May (autumn) 2019, relating them to possible ocean warming events. The thermal anomalies charts were taken by INMET and the fieldwork were developed based on the AGRRA Protocol. Two 10 m long transects were used, placed continuously and parallel to the larger rocky shore axis, where quadrats of 36X36 cm were distributed at positions demarcated in 1, 3, 5, 7 and 9 m along these transects and photographed to obtain the data. The total number of colonies, number of bleached colonies and the average area of each species of benthic cnidarians affected by different bleaching intensities (strong degree and weak degree) were calculated. Four species of benthic cnidarian have been identified, among them the Millepora alcicornis and the Palythoa caribaeorum were the ones that suffer bleaching (strong degree and weak degree), with an increase mainly of the strong degree in the month of May - a period preceded by a thermal anomaly of 2.0 ºC throughout the month of March and in the second half of April; while the Siderastrea stellata and Zoanthus sp. did not suffer bleaching.
Keywords: Coral bleaching, Thermal anomalies, Forno Beach, Arraial do Cabo, Rocky shore.
Resumen: El presente estudio tiene como objetivo describir casos de blanqueamiento de corales y otros cnidarios bentónicos en la orilla rocosa de la Playa de Forno (Arraial do Cabo, Brasil), observadas en febrero (verano) y mayo (otoño) de 2019, relacionándolas con posibles eventos de calentamiento de las aguas superficiales del océano. Los mapas de anomalías térmicas fueron obtenidos por INMET y el trabajo de campo se desarrolló con base en el Protocolo AGRRA. Se utilizaron dos transectos de 10 m de largo, paralelo al eje mayor de la orilla rocosa, donde se distribuyeron cuadrados de 36x36 cm, en las posiciones demarcadas em 1, 3, 5, 7 y 9 m a lo largo de estos transectos y cuadrados fotografiados para obtener los datos. Se calculó lo siguiente: el número total de colonias, el número de colonias blanqueadas y el área media de cada especie de cnidario bentónico afectada por diferentes intensidades de blanqueamiento (débil o fuerte). Se han identificado cuatro especies de cnidarios bentónicos, entre estas Millepora alcicornis e Palythoa caribaeorum sufrieron blanqueamiento fuerte y débil, con aumento principalmente de blanqueamiento fuerte en mayo - período precedido por una anomalía térmica de 2.0 ºC durante todo el mes de marzo y en la segunda quincena de abril; mientras que Siderastrea stellata y Zoanthus sp. no han sido blanqueadas.
Palabras clave: Blanqueamiento de coral, Anomalías térmicas, Playa de Forno, Arraial do Cabo, Orilla rocosa.
1 Introdução
Os ecossistemas recifais são de grande importância ecológica, pois atuam como áreas de desova, abrigo, reprodução e alimentação para uma grande variedade de organismos marinhos, além de servirem como uma barreira mitigadora da erosão costeira (MOBERG; FOLKE, 1999). Economicamente, geram inúmeros bens e serviços ecossistêmicos utilizados pela sociedade (CASTRO; ZILBERBERG, 2016; PICCIANI et al., 2016). Destacam-se pela grande riqueza e abundância de organismos, que produzem uma complexa comunidade biológica de considerável beleza cênica, o que torna os ecossistemas recifais pontos turísticos relevantes no mundo todo, gerando empregos e fontes de renda diretos e indiretos (CASTRO; ZILBERBERG, 2016; PICCIANI et al., 2016).
Distribuídos ao longo de 3.000 km da costa brasileira, os ecossistemas recifais ocorrem em latitudes equatoriais até a subtropicais, compreendendo a maior e mais rica área do Atlântico Sul (LEÃO et al., 2016). Apesar de serem considerados menos diversos, quando comparados a outros ambientes recifais distribuídos no mundo, os recifes brasileiros, que possuem 23 espécies de corais e cinco espécies de hidrocorais (LEÃO et al., 2016), apresentam alto grau de endemismo (LEÃO; KIKUCHI; TESTA, 2003). Neste contexto, chama-se a atenção para o fato de que seis das espécies de corais construtores dos recifes brasileiros são endêmicas: Mussismilia braziliensis, Mussismilia hispida, Mussismilia hartii, Favia leptophylla, Siderastrea stellata e Favia gravida (LEÃO; KIKUCHI; TESTA, 2003).
Em relação aos impactos, estima-se que as principais causas da degradação dos ambientes recifais atualmente ocorrem em função do amplo desenvolvimento costeiro de forma desordenada, o que leva a outros desdobramentos como: aumento da poluição, aumento da sedimentação e consequente soterramento dos organismos e a sobre-exploração dos recursos marinhos (FERREIRA; MAIDA, 2006; LEÃO et al., 2016). Destacam-se também outros impactos globais, como o aquecimento e acidificação dos oceanos (BRIDGE; HUGUES; BONGAERTS, 2013), que levam a anomalias térmicas das águas superficiais, que causam o fenômeno do branqueamento. Além das anomalias térmicas, os corais também são afetados diretamente pelos mergulhos recreativos que, na maioria das vezes, ocorre de maneira inadequada permitindo o contato físico com os organismos, promovendo sua quebra, além da ressuspensão de sedimento, deslocamento das colônias (GUZNER et al., 2010; LUNA-PÉREZ; VALLE-PÉREZ; SÁNCHEZ-LIZASO, 2011; POONIAN; DAVIS; McNAUGHTON, 2010) e danos causados pelo ancoramento das embarcações (DINSDALE; HARRIOTT, 2004).
O branqueamento consiste na perda da coloração dos corais vivos devido à expulsão dos seus endossimbiontes (zooxantelas), e/ou de seus pigmentos fotossintéticos (KELMO, 1998). As zooxantelas são responsáveis por produzir componentes orgânicos, através da fotossíntese, que lhes provê assim como aos corais, em relação mutualística. Em contrapartida, o coral provê abrigo para as algas e lhes fornece elementos químicos necessários à sua sobrevivência (HISSA et al., 2009; LEÃO; KIKUCHI; OLIVEIRA, 2008). Ainda segundo Kelmo (1998), é possível destacar outros efeitos oriundos desse processo de branqueamento, tais como: aumento na taxa respiratória dos corais e o declínio nos níveis de proteínas, lipídios e carboidratos das colônias escleractínias. Além disso, como constatou Correia (2016), muitos eventos de doenças são identificados durante, ou após episódios de branqueamento, uma vez que colônias branqueadas são mais suscetíveis a infecções por terem as propriedades antibióticas do muco reduzidas.
Outros cnidários bentônicos como os zoantídeos (Palythoa e Zoanthus, por exemplo), também são acometidos pelo branqueamento devido às anomalias térmicas (KEMP et al., 2006; LESSER et al., 1990; SANTOS et al., 2016). Esses cnidários não secretam o esqueleto calcário como nos corais, entretanto são frequentemente os principais organismos que formam a estrutura das comunidades bentônicas de ambientes recifais nas regiões tropicais e temperadas (CASTRO; PIRES, 2001; OIGMAN-PSZCZOL; FIGUEIREDO; CREED, 2004).
O fenômeno do branqueamento de corais vem se intensificando em episódios de larga escala e maior tempo de duração (HUGHES et al., 2019). Hughes et al. (2018) verificaram que a frequência e a intensidade de eventos de branqueamento estão ocorrendo de forma global e insustentável. Em 1998, no Oceano Índico mais de 90% dos recifes de corais morreram em decorrência, principalmente, da elevação da temperatura das águas superficiais (HOEGH-GULDBERG, 1999). A frequência de eventos de branqueamento severo quintuplicou nas últimas quatro décadas, de um a cada 25 a 30 anos no início dos anos 1980, a um a cada 5,9 anos, em 2016 (HUGHES et al., 2018). Devido ao aquecimento global se observa o gradual aumento das temperaturas médias das águas superficiais dos oceanos nas regiões tropicais, principalmente quando ocorre o fenômeno do El-Niño (ENSO) (HUGHES et al., 2018).
No Brasil, o fenômeno de branqueamento tem sido registrado desde 1993, conforme a tendência global (CASTRO, PIRES, 1999; LEÃO et al., 2010; SOUZA, 2013). As anomalias térmicas são uma das causas deste fenômeno e são monitoradas, na costa brasileira, por imagens de satélites do National Oceanic and Atmospheric Administration (NOAA), desde 1998 (MIRANDA; CRUZ; LEÃO, 2013). Conforme estudo realizado por Leão, Kikuchi e Oliveira (2008), entre os anos de 1998 a 2008, é visível a ocorrência de recifes de corais branqueados na costa do estado da Bahia, variando em diferentes graus. Dentre as regiões afetadas segundo a pesquisa, destacam-se: Litoral Norte, Baía de Todos os Santos, ilhas de Tinharé e Boipeba, Cabrália, Itacolomis e Abrolhos (LEÃO; KIKUCHI; OLIVEIRA, 2008).
Em relação a outros ambientes recifais no mundo, Mies et al. (2020) verificaram que os recifes da região do Atlântico Sul apresentaram proporcionalmente menos episódios de branqueamento de corais com mortalidade do que nas regiões do Indo-Pacífico e do Caribe. A maioria dos episódios de branqueamento ocorridos no Atlântico Sul ocorreram no nordeste do Brasil, particularmente na costa do estado da Bahia (MIES et al., 2020). No entanto, em 80% dos episódios de branqueamento não foi observada a mortalidade dos corais nesta região, dados que corroboraram trabalhos de Leão et al. (2016) e Teixeira et al. (2019). Esse fato pode estar relacionado à maior resistência a condições adversas, por esses corais, que geralmente ocupam zonas de maior profundidade, serem tolerantes a maior turbidez e concentração de nitrato, mais resistentes morfologicamente (maioria massivos com tecidos mais espessos) e apresentarem maior diversidade de algas simbiontes (MIES et al., 2020), o que os torna relativamente menos afetados pelo branqueamento.
Em relação à distribuição dos ambientes recifais no Brasil, o município de Arraial do Cabo, na região de Cabo Frio (RJ) é considerada o limite sul, para a ocorrência de várias espécies de corais da costa brasileira (CASTRO; PIRES, 2001; LIMA; COUTINHO, 2016; LABOREL, 1970). Seu território é reconhecidamente rico em fauna e flora marinha e conta com a Reserva Extrativista Marinha para conservar os interesses ecológico-sociais, e para proteger a cultura da pesca artesanal em Arraial do Cabo (FONSECA-KRUEL; PEIXOTO, 2004). Nesta região, a Praia do Forno, considerada um dos principais atrativos turísticos, é uma das que mais recebe visitantes e, portanto, merece uma especial atenção, pois abriga uma grande variedade de peixes, dentre outras espécies marinhas, por conter áreas protegidas e locais sujeitos à exposição de ondas (FERREIRA; GONÇALVES; COUTINHO, 2001).
Em Arraial do Cabo, a prática da sobrepesca, a introdução de espécies exóticas e de agentes patogênicos de diferentes regiões do mundo (MELO et al., 2009), somado ao turismo predatório com grandes massas se deslocando todos os anos para esta região (CARNEIRO; VIEIRA; COLUCCI, 2012) são as principais causas da degradação dos ecossistemas marinhos locais. Com isto, as atividades de turismo, e do Porto do Forno oferecem altos riscos antropogênicos, tornando a sustentabilidade da Reserva Extrativista um desafio constante (CARNEIRO; VIEIRA; COLUCCI, 2012). Além disso, a região é considerada a capital brasileira do mergulho, e, portanto, a saúde dos corais e de outros cnidários bentônicos é de vital importância para a manutenção das atividades econômicas da população residente.
Sendo assim, em função da grande importância ecológica e econômica desses organismos, o objetivo geral deste estudo é descrever as ocorrências de branqueamento de corais e demais cnidários bentônicos presentes no costão da Praia do Forno (Arraial do Cabo, RJ), observadas no mês de fevereiro (verão) e maio (outono) do ano de 2019, relacionando-as com possíveis eventos de aquecimento das águas superficiais do oceano.
2 Metodologia
2.1 Área de estudo
O presente estudo foi realizado em Arraial do Cabo, RJ, costa sudeste do Brasil (22º 56’ 06.0”S e 42° 00’ 56.1”W), onde ocorre o fenômeno da ressurgência (VALENTIN, 1984). Esse fenômeno consiste no afloramento das Águas Centrais do Atlântico Sul (ACAS) que são frias e ricas em nutrientes e que, portanto, fertilizam a cadeia trófica no encontro com as correntes tropicais, tornando a região mais rica e com abundante biodiversidade (CARNEIRO; VIEIRA; COLUCCI, 2012).
A coleta de dados foi realizada na Praia do Forno (Figura 1), que apresenta uma extensão de 580 m. O costão rochoso visitado é caracterizado por uma estrutura de rochas graníticas, terminando em um fundo de areia, com a presença de cascalho em alguns pontos (FERREIRA; GONÇALVES; COUTINHO, 2001). Essa enseada proporciona condições para o desenvolvimento de uma rica comunidade recifal (FERREIRA; GONÇALVES; COUTINHO, 2001; GUIMARAENS; COUTINHO, 1996; ORNELLAS; COUTINHO, 1998), sendo associada a áreas mais rasas, entre 2 e 3 m de profundidade, e temperatura da água com média maior do que as áreas expostas ao oceano (aproximadamente 22 ºC) (FERREIRA; GONÇALVES; COUTINHO, 2001).
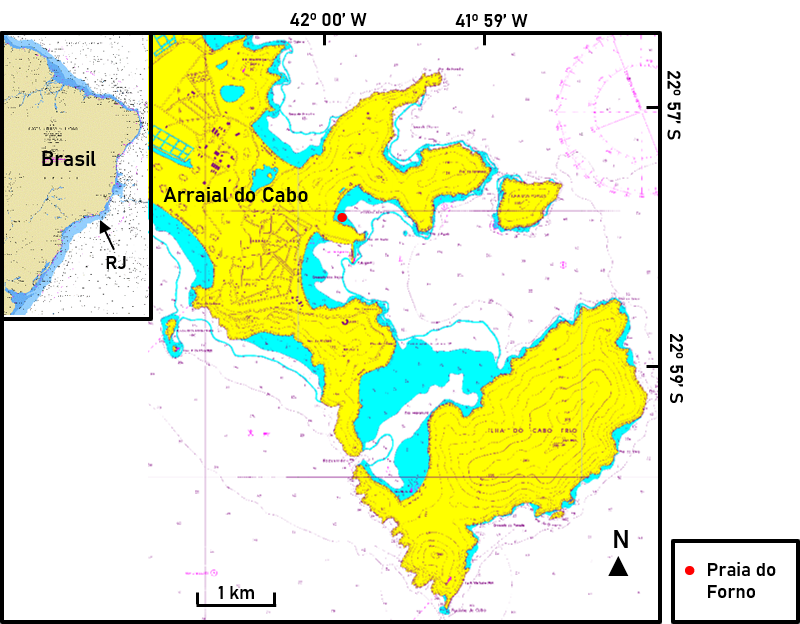
Figura 1.
Localização da Praia do Forno em Arraial do Cabo/RJ e a área analisada durante o presente estudo
Fonte: adaptado de https://www.marinha.mil.br/chm/dados-do-segnav/cartas-raster
As expedições foram realizadas nos dias 22 de fevereiro e 06 de maio de 2019, através de mergulho livre. Nas visitas à campo, foram levantados dados de branqueamento classificados como forte e fraco, levando-se em consideração a intensidade de coloração das colônias (branca e pálida, respectivamente).
2.2 Levantamento dos dados climáticos
As anomalias térmicas para a região da Praia do Forno foram obtidas utilizando-se as cartas de anomalias térmicas da superfície do mar produzidas pelo Instituto Nacional de Meteorologia (INMET), disponíveis no seguinte portal eletrônico: http://www.inmet.gov.br/portal/index.php?r=clima/temperaturaSuperficieMar. Foram utilizados dados da 1.ª e 2.ª quinzena de cada mês no período entre janeiro e maio de 2019, os quais correspondem ao final do verão e ao início do outono no hemisfério sul, respectivamente.
2.3 Aquisição dos dados de campo
Ambos os trabalhos de campo foram desenvolvidos com adaptações da metodologia proposta no Protocolo AGRRA (Atlantic and Gulf Rapid Reef Assessment, LANG et al., 2010). A técnica utilizada para a coleta dos dados descrita no protocolo AGRRA baseia-se na utilização de senso visual dos organismos, por meio de unidades amostrais quadradas dispostas ao longo de transectos. Para avaliar as condições dos corais no único local estudado foram colocados de forma contínua dois transectos de 10 m de comprimento, distribuídos paralelamente ao eixo maior do infralitoral do costão rochoso. Ao longo de cada transecto, foram distribuídos cinco quadrados (36 cm x 36 cm) nas posições demarcadas em 1, 3, 5, 7 e 9 m para a obtenção dos dados. Os transectos não foram fixados, mas procurou-se posicioná-los no mesmo local durante as duas expedições. Cada quadrado foi fotografado, para a identificação de possível branqueamento dos cnidários. No dia 22 de fevereiro, foi utilizada a câmera de alta resolução do IPHONE 7, dentro de uma capa a prova d’água universal, e no dia 6 de maio, foram utilizadas as câmeras de alta resolução do IPHONE 7 dentro de uma capa impermeável, e do modelo Samsung S860 em caixa estanque.
A análise dos dados foi realizada adaptando a metodologia descrita por Miranda, Cruz e Leão (2013). Desta forma, foram obtidos dados baseados nas fotografias dos quadrados feitas durante as expedições (Figura 2). Em cada um destes foram contados o número de colônias (NC), número de colônias branqueadas (NCB) e a média (DP) da porcentagem da área estimada da colônia afetada por diferentes intensidades de branqueamento (fraco ou forte) por cada espécie de organismo. Foi realizado o cálculo estimado da porcentagem da área afetada da colônia pelo branqueamento total (BT), o branqueamento fraco (BFR) e o branqueamento forte (BFO).
A normalidade dos dados foi testada utilizando-se o teste de Skewness e como parte dos dados não foram paramétricos, optamos por avaliar possíveis diferenças estatísticas entre os parâmetros por meio do teste de Kruskal-Wallis com nível de significância p<0,05 que indica diferença significativa entre os parâmetros analisados (ZAR, 1999). Comparamos as médias dos parâmetros do branqueamento total (BT), branqueamento fraco (BFR) e branqueamento forte (BFO) entre as duas expedições e avaliamos as diferenças entre as médias dentro da expedição de maio (período que foi antecedido pelos maiores valores de anomalia térmica) do branqueamento total (BT), branqueamento fraco (BFR) e branqueamento forte (BFO) entre as diferentes espécies encontradas.
Foram realizadas correlações entre: a temperatura da água da superfície do mar (TSM) e a porcentagem do branqueamento total (BT) – (TSM x BT), entre esta e o branqueamento fraco (BFR) – (TSM x BFR), e entre esta e o branqueamento forte (BFO) – (TSM x BFO), respectivamente para as duas espécies que apresentaram branqueamento. Avaliamos em qual correlação o coeficiente se apresentou positivo através do sinal â1 da regressão (o que indica uma relação de proporcionalidade direta entre as variáveis) e a qualidade da regressão através do valor absoluto de r (ARANGO, 2005).

Figura 2.
Quadrats utilizado para aquisição das fotos subaquáticas dos cnidários bentônicos na Praia do Forno em Arraial do Cabo/RJ
Fonte: os autores
A intensidade do branqueamento (fraco ou forte) foi atribuída de acordo com a tonalidade da colônia (pálida ou branca) (LEÃO; KIKUCHI; OLIVEIRA, 2008).
3 Resultados
Foram identificadas quatro espécies de cnidários bentônicos no costão rochoso estudado: o hidrocoral Millepora alcicornis, o coral escleratíneo Siderastrea stellata e as duas espécies de zoantídeos Palythoa caribaeorum e Zoanthus sp.
3.1 Relação entre anomalias térmicas e intensidades de branqueamento
Pôde-se observar que, em janeiro, a anomalia térmica manteve-se constante em 1,0 °C nas duas quinzenas, apresentando queda de 0,5 °C na primeira quinzena de fevereiro, e retornando ao valor de 1,0 °C na segunda quinzena do mesmo mês. Posteriormente, o valor de anomalia térmica observada foi de 1,0 °C durante a primeira quinzena de abril e na segunda quinzena de maio. Os valores máximos de anomalia (2,0 °C), foram observados ao longo de março, na segunda quinzena de abril e na primeira de maio (Figura 3).
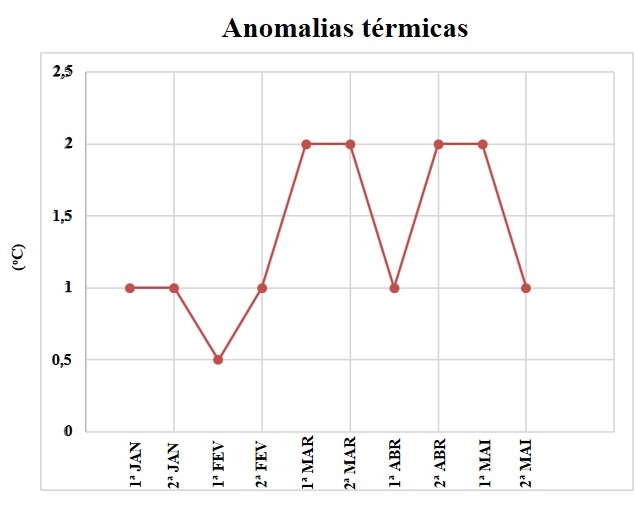
Figura 3.
Anomalia da temperatura das águas superficiais oceânicas para a costa de Arraial do Cabo em 2019, obtidas de imagens orbitais. Origem dos dados http://www.inmet.gov.br/portal/index.php?r=clima/temperatura SuperficieMar. O tempo cronológico utilizado são as 1ª e 2ª quinzenas dos cinco (5) meses apresentados: janeiro, fevereiro, março, abril e maio.
Fonte: os autores
Quanto ao fenômeno do branqueamento, nos dois meses de estudo (fevereiro e maio), registrou-se a presença de branqueamento forte e fraco na área da coleta de dados. Do total de quatro espécies de cnidários bentônicos registradas na Praia do Forno, em duas delas foi identificada presença de algum grau de branqueamento, são elas Palythoa caribaeorum e Millepora alcicornis.
A anomalia térmica na primeira quinzena de fevereiro foi de 0,5 °C e na segunda quinzena deste mês foi de 1,0 ºC (Figura 3), durante essa primeira expedição foi registrado branqueamento em 76% das 25 colônias avaliadas nesse período (Tabela 1). Anomalias térmicas ocorreram entre os meses de março até o período da primeira quinzena de maio, quando realizamos a segunda expedição. A anomalia térmica nas duas quinzenas do mês de março atingiu valor de 2,0 °C, assim como na segunda quinzena de abril e na primeira quinzena de maio. Durante essa segunda expedição, foi registrado branqueamento em 66,6% das 27 colônias avaliadas nesse período.
3.2 Branqueamento das espécies avaliadas
As espécies Millepora alcicornis e Palythoa caribaeorum (Figura 4) apresentaram diferentes porcentagens e intensidades (forte/fraco) de branqueamento nas áreas estudadas (Tabela 1). Zoanthus sp. e Siderastrea stellata (Figura 4) não sofreram branqueamento.

Figura 4.
Espécies de cnidários bentônicos analisadas na Praia do Forno, Arraial do Cabo/RJ. A - Palythoa caribaeorum; B -Zoanthus sp; C - Millepora alcicornis; D - Siderastrea stellata; E - Millepora alcicornis com branqueamento forte; F - Palythoa caribaeorum com branqueamento forte; G - Millepora alcicornis com branqueamento fraco; H - Palythoa caribaeorum com branqueamento fraco.
Fonte: os autores
Palythoa caribaeorum – foi a espécie de cnidário bentônico mais abundante. Do total de colônias observadas, 76,20% foram afetadas por branqueamento durante a primeira expedição, em que ocorreu uma anomalia térmica de 1,0 °C (Figura 3). O branqueamento afetou 35,71% da área colonial deste zoantídeo na primeira expedição, destes 22,13% foi por branqueamento fraco e 13,59% foi por branqueamento forte (Tabela 1). Na segunda expedição, o branqueamento afetou 87,5% do número total de colônias observadas quando ocorreu uma anomalia de 2,0 °C durante a segunda quinzena de abril e primeira quinzena de maio (Figura 3). A área colonial afetada por branqueamento foi de 49,07% - destes 18,59% foi por branqueamento fraco e 30,47% foi por branqueamento forte (Tabela 1). O teste de Kruskal-Wallis não mostrou diferença significativa entre a primeira expedição e a segunda expedição para os parâmetros branqueamento total (H =29,00; p > 0,05), branqueamento fraco (H = 42,50; p > 0,05) e branqueamento forte (H = 42,50; p > 0,05).
Millepora alcicornis – foi a segunda espécie de cnidário bentônico mais abundante e a mais afetada pelo branqueamento forte durante as duas expedições, ambas com 100% do número total das colônias observadas apresentando algum tipo de branqueamento (fraco ou forte). O branqueamento afetou 21% da área colonial deste hidrocoral na primeira expedição, sendo 5% por branqueamento fraco e 15,90% por branqueamento forte (Tabela 1). Na segunda expedição, 51,22% da área colonial deste cnidário apresentou branqueamento, sendo o mais alto valor observado entre todas as espécies estudadas, além de ter o mais alto percentual de branqueamento forte (41,65%); o branqueamento fraco apresentou 9,52% da área colonial desta espécie (Tabela 1). O teste de Kruskal-Wallis não mostrou diferença significativa entre as duas expedições para os parâmetros branqueamento total (H = 4,50; p > 0,05), branqueamento fraco (H = 3,00; p > 0,05) e branqueamento forte (H = 4,00; p > 0,05).
Comparando estatisticamente as quatro espécies observadas na expedição de maio de 2019, o teste de Kruskal-Wallis não mostrou diferença significativa para os parâmetros branqueamento total (H = 7,77; p > 0,05), branqueamento fraco (H = 3,46; p > 0,05) e o branqueamento forte (H = 7,50; p > 0,05). Embora os testes não tenham mostrado uma diferença significativa de branqueamento relacionado com a anomalia térmica de 2 ºC no período correspondente à expedição de maio, há uma tendência no aumento principalmente das proporções de branqueamento forte (BFO) nas espécies que sofreram algum grau de branqueamento (Tabela 1).
Foi feita a análise de correlação entre a temperatura da superfície do mar (TSM) e a porcentagem do branqueamento total (BT), branqueamento fraco (BFR) e branqueamento forte (BFO) das duas espécies que apresentaram branqueamento (Millepora alcicornis e Palythoa caribaeorum). Os resultados mostraram que somente a correlação entre a temperatura da superfície do mar (TSM) e o branqueamento forte (BFO) da espécie Palythoa caribaeorum foi positiva e significativa (r=0,61). Apesar de essa correlação apresentar-se fraca, indica uma tendência de influência da temperatura da superfície do mar com o branqueamento forte nesta espécie.
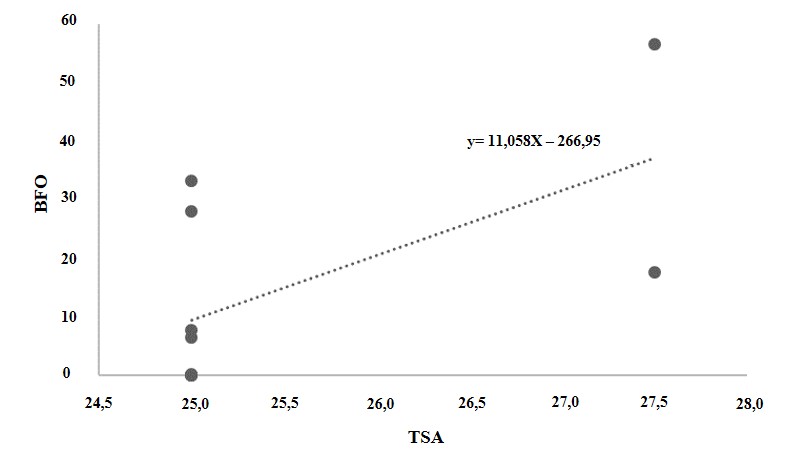
Figura 5.
Correlação entre a temperatura superficial do mar (TSM) X branqueamento forte (BFO) sofrido pela espécie Palythoa caribaeorum da Praia do Forno, Arraial do Cabo em 2019 (r = 0,61). Origem dos dados http://www.inmet.gov.br/portal/index.php?r=clima/temperatura SuperficieMar.
Legenda: BFO – branqueamento forte; TSM – temperatura da superfície do mar
Fonte: os autores
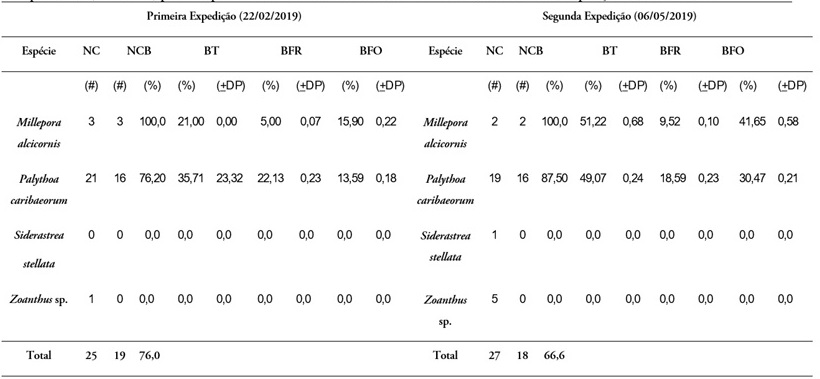
Fonte: Os autoresLegenda: NC - número de colônias; NCB - número de colônias branqueadas; BT - branqueamento total; BFR - branqueamento fraco; BFO - branqueamento forte; DP - desvio padrão.
4 Discussão
No presente trabalho foram identificadas quatro espécies de cnidários bentônicos (Millepora alcicornis, Siderastrea stellata, Palythoa caribaeorum e Zoanthus sp.). Em trabalho realizado por Rogers et al. (2014) nesta mesma área de estudo, foi identificada também a espécie Mussismiliahispida. Em contrapartida, espécimes do gênero Zoanthus foram registradas no presente estudo, corroborando com o realizado por Macedo (1986), nesta praia. Quanto à Palythoa caribaeorum esta é uma espécie frequentemente encontrada cobrindo costões rochosos da costa sudeste do Brasil (CASTRO; PIRES, 2001; OIGMAN-PSZCZOL; CREED, 2004), e também recifes biogênicos na costa nordeste (RAMOS et al., 2010), e, portanto, o seu registro já era esperado para a região estudada. Em relação à Millepora alcicornis,Leão, Kikushi e Oliveira (2008) verificaram que esta também foi a segunda espécie de coral mais abundante em recifes costeiros da Bahia, assim como observado na Praia do Forno (Arraial do Cabo, RJ), neste estudo.
O branqueamento de P. caribaeorum e M. alcicornis pode estar relacionado com a ocorrência de anomalias térmicas das águas superficiais do oceano, conforme observado. O registro de uma anomalia térmica de 2,0 °C que perdurou durante quatro semanas (entre abril e maio) pode ter provocado a maior proporção de branqueamento forte, registrada neste estudo. Essa hipótese é reforçada pelo estudo realizado por Leão, Kikuchi e Oliveira (2008), que observaram que anomalias térmicas entre 0,50 °C e 0,75 °C, durante três semanas causaram o branqueamento de 20% recifes de corais registrados no mês de maio de 2005, no arquipélago de Abrolhos. Isso sugere que uma anomalia de 2,0 °C durante um período de quatro semanas pode ser considerado suficiente para causar branqueamento forte em alguns dos cnidários bentônicos da Praia do Forno.
Os recifes de corais da região da costa do nordeste do Brasil é onde tem ocorrido a maioria dos episódios de branqueamento do Atlântico Sul, principalmente no litoral do estado da Bahia. A mortalidade de corais em decorrência do processo de branqueamento na região do oceano Atlântico Sul foi menor do que 20% (LEÃO et al., 2016; MIES et al., 2020; TEIXEIRA et al., 2019), o que é aproximadamente 60% menor que no oceano Indo-Pacífico (MIES et al., 2020; RODGERS et al., 2017) e 50% menor que a que ocorre na região do Caribe (MIES et al., 2020). A maior tolerância dos corais do Atlântico Sul deve-se à adaptação aos estressores ambientais como maior turbidez e concentração de nitrato; apresentam em sua maioria forma massiva com tecidos mais espessos, possuem maior diversidade de endossimbiontes e habitam zonas de maior profundidade – fatores que em combinação tornaram as espécies mais resistentes ao branqueamento e mortalidade (MIES et al., 2020). Registrar e compreender a recuperação em recifes marginais (ou seja, em condições subótimas por exemplo de turbidez, baixa temperatura) pode gerar conhecimentos importantes sobre o branqueamento como um fenômeno global (LONGO; CORREIA; MELLO, 2020).
Em relação à suscetibilidade e ao grau de branqueamento, foi observado no presente estudo, que a espécie Palythoa caribaeorum foi a mais abundante (maior área colonial) e a segunda mais intensamente afetada por este fenômeno, enquanto Milleporaalcicornis foi proporcionalmente mais branqueada. Rogers et al. (2014) também indicaram a maior abundância de Palythoa caribaeorum seguida por Millepora alcicornis – resultados semelhantes foram encontrados no presente trabalho. Em estudo realizado por Hissa et al. (2009) foi observado que P. caribaeorum apresentou sensibilidade intermediária a temperaturas mais altas, uma vez que houve a ocorrência de branqueamento completo em 48 horas, quando submetidos a 45 °C e em 72 horas, quando submetidos a 40 °C. Já a 35 °C, as colônias não apresentaram branqueamento (HISSA et al., 2009).
Em relação à M. alcicornis, esta espécie apresentou proporção de branqueamento do grau forte maior que no mês de fevereiro, além de apresentar maior proporção de branqueamento forte, quando comparada com P. caribaeorum. Esses resultados corroboram com a pesquisa de Miranda, Cruz e Leão (2013), que também trabalharam com Millepora alcicornis, nos recifes de Caramuanas, no estado da Bahia. Os autores observaram que essa foi a espécie mais afetada pelo branqueamento forte, durante as anomalias térmicas, tendo 37,14% de sua área total afetada. No presente trabalho e no publicado por Miranda, Cruz e Leão (2013) esta espécie mostrou-se sensível ao branqueamento, mas sem taxa de mortalidade relacionada ao evento da anomalia térmica. Duarte et al. (2020) analisando M. alcicornis na Reserva Extrativista Marinha de Corumbau (litoral do estado da Bahia) registraram mortalidade entre 43% e 89% entre os lugares amostrados, coincidindo com a onda de calor registrada no ano de 2019.
Embora os testes não tenham mostrado diferença significativa do branqueamento relacionado à anomalia térmica no período correspondente à anomalia térmica de 2 °C (segunda quinzena de abril - primeira quinzena de maio) há uma tendência de influência da temperatura da superfície do mar com a ocorrência de branqueamento ocorrido nas duas espécies afetadas (P. caribaeorum, M. alcicornis). Importante salientar a necessidade de estudos futuros aumentando a amostragem para se confirmar ou não essa tendência da maior proporção de branqueamento relacionado com as anomalias térmicas e se o fenômeno da ressurgência pode ter relação com a menor porcentagem de branqueamento, devido à habilidade dos corais em se adaptar às condições (baixa turbidez e temperatura) onde ocorrem esse fenômeno (LIMA; COUTINHO, 2016).
Em relação à resistência ao fenômeno do branqueamento, foi observado que Zoanthus sp. e Siderastreastellata não sofreram esse impacto, nesse período, assim como observado por Rogers et al. (2014), que detectaram uma baixa porcentagem de branqueamento na espécie S. stellata (0,2%) na Praia do Forno. A partir das anomalias térmicas baixas de fevereiro (de até 1,0 °C), a espécie Palythoa caribaeorum apresentou grau de branqueamento predominante fraco, enquanto no mês de maio, quando as anomalias térmicas encontravam-se relativamente altas (2,0 °C), as duas espécies (Palythoa caribaeorum e Millepora alcicornis) foram afetadas pelo branqueamento forte, sendo que a Milleporaalcicornis apresentou uma proporção maior, com 100% de suas colônias branqueadas - com 41,65% de sua área afetada pelo branqueamento forte.
De acordo com o estudo de Macedo (1986), na Praia do Forno, Arraial do Cabo/RJ, foi encontrada grande predominância de zoantídeos nos costões rochosos próximos à praia (Palythoa caribaeorum e Zoanthus sp.). Através das proporções e porcentagem de branqueamento, pode-se observar que a espécie Zoanthus sp. foi a mais resistente, levando-se em consideração que nenhuma colônia sofreu o branqueamento, em nenhum dos meses avaliados. Desse modo, é possível afirmar que a presente espécie é capaz de suportar as anomalias térmicas de até 2,0 °C nas condições ambientais presentes no local de estudo. Esse fato coincide com o relato de Hissa et al. (2009), cujas amostras de Zoanthussociatus coletadas nos recifes de arenito em duas praias na costa cearense (Paracuru e Praia dos Dois Coqueiros) mostraram-se mais resistentes à elevação de temperatura a partir de 40 ºC.
A espécie Siderastrea stellata também não sofreu branqueamento, sugerindo que também é capaz de suportar anomalias térmicas de até 2,0 °C nas condições ambientais presentes no local de estudo. No presente estudo não fizemos a caracterização da zooxantela presente nos corais e zoantídeos. Um fator que contribui com a maior resistência e menor porcentagem de branqueamento em S. stellata é a simbiose com a zooxantela Cladocopium sp., que apresenta uma ampla tolerância a temperatura (KARAKO-LAMPERT et al., 2004), presença esta corroborada por Sassi et al. (2014) em pesquisa realizada nesses escleratíneos dos recifes costeiros de Cabo Branco (litoral do estado da Paraíba).
Para finalizar, é necessário salientar que estudos de monitoramento desses organismos na área de estudo são extremamente importantes, pois os eventos de branqueamento em ambientes recifais tem como consequência, direta e indireta, a perda de cobertura da área recifal; declínio da riqueza de espécies associadas de forma geral (algas, vertebrados, invertebrados); dominância de espécies resistentes ao branqueamento e consequente perda de diversidade (BURT; AL-HARTHI; AL-CIBAHI, 2011; GLYNN, 1993); desestabilização da estrutura dos corais com danos em sua estrutura e que, dependendo da escala e magnitude, a recuperação pode ser um processo muito lento (GLYNN, 1993). Portanto, o manejo dos ambientes recifais ao redor do mundo é de grande importância para a preservação da diversidade biológica marinha, que garantem inúmeros serviços ecossistêmicos para sociedade (CASTRO; ZILBERBERG, 2016; MOBERG; FOLKE, 1999; PICCIANI et al., 2016).
5 Conclusões
As quatro espécies de cnidários registradas já eram previstas para a área de estudo, e apresentaram diferentes respostas de branqueamento, em relação à espécie, e em relação ao tempo de estudo, relacionado diretamente à variação térmica das águas superficiais. Desta forma, pôde ser observado que as espécies mais sensíveis às anomalias térmicas registradas foram Palythoa caribaeorum e a Millepora alcicornis, enquanto que Zoanthus sp. e a Siderastrea stellata foram mais resistentes. Foi observada, portanto, uma tendência da influência da temperatura da superfície do mar com a ocorrência de branqueamento ocorrido nessas duas espécies, e que embora não estatisticamente significativa, aponta para a necessidade de uma amostragem temporal mais extensa, para a melhor compreensão do branqueamento na região de Arraial do Cabo, área de intensa influência da ressurgência.
Em vista as causas da degradação dos corais, é fundamental a adoção de medidas mitigadoras para esse processo. Fatores antropogênicos que influenciam o aumento das anomalias térmicas das águas superficiais dos oceanos e os distúrbios do El Niño, devido às mudanças climáticas globais estão associados ao branqueamento dos corais. Esse aumento da temperatura das águas superficiais dos oceanos está ligado principalmente com a queima de combustíveis fósseis e, consequentemente, a liberação do gás carbônico, um dos principais fatores do efeito estufa. Além das mudanças na matriz energética de maneira global, outras medidas importantes para a preservação dos corais devem ser tomadas para diminuir a poluição, sedimentação, sobre-explotação e o turismo destrutivo, juntamente com a conscientização por mudanças de hábito do cidadão comum.
Agradecimentos
Agradecemos a Escola Técnica Estadual Helber Vignoli Muniz (FAETEC), pois, este manuscrito é resultado do Trabalho de Conclusão do Curso Técnico Integrado em Meio Ambiente; somos gratos a todo o suporte que nos foi oferecido. Agradecemos também aos editores e aos revisores anônimos que contribuíram com valiosas sugestões aperfeiçoando a versão final deste artigo.
Referências
ARANGO, H. G. Bioestatística: Teórica e computacional. Rio de Janeiro: Guanabara Koogan, 2005. 423p.
BRIDGE, T. C. L. et al. Call to protect all coral reefs. Nature climate Change, v.3, p. 528-530, 2013.
BURT, J.; AL-HARTHI, S.; AL-CIBAHY, A. Long-term impacts of coral bleaching events on the world’s warmest reefs. Marine Environmental Research, v. 72, p. 225-229, 2011.
CAMPOS, F. F. Diversidade de bactérias associadas ao muco do zoantídeo Palythoa Caribaeorum (Cnidaria, Anthozoa) do litoral sul de Pernambuco. 2011. 73 f. Dissertação (Mestrado em Saúde Humana e Meio Ambiente) - Universidade Federal de Pernambuco, Recife, 2011.
CARNEIRO, A. M.; VIEIRA, L.; COLUCCI, A. Projeto ressurgência: Extensão participativa de SMS para pesca artesanal na Reserva Extrativista Marinha de Arraial do Cabo, RJ. Revista Brasileira de Ergonomia, v. 7, n. 1, p. 55-73, 2012.
CASTRO, C. B.; PIRES, D. O. A bleaching event on a Brazilian coral reef. Revista Brasileira de Oceanografia, v. 47, n. l, p. 87-90, 1999.
CASTRO, C. B.; PIRES, D. O. Brazilian coral reefs: what we already know and what is still missing. Bulletin of Marine Science, v. 69, n.2, p. 357–371, 2001.
CASTRO, C. B.; ZILBERBERG, C. Recifes brasileiros, sua importância e conservação. In: ZILBERBERG, C. et al. Conhecendo os Recifes Brasileiros: Rede de Pesquisas Coral Vivo. Rio de Janeiro: Museu Nacional, Universidade Federal do Rio de Janeiro, 2016. Cap. 1.
CORREIA, L. F. C. Padrão de branqueamento de Siderastrea stellata (Verril, 1868) em dois complexos recifais areníticos do Atlântico Sul. 2016. 36 f. Monografia (Graduação em Ecologia – Bacharelado) - Universidade Federal do Rio Grande do Norte, Natal, 2016.
DINSDALE, E.A.; HARRIOTT, V. J. Assessing anchor damage on coral reefs: A case study in selection of environmental indicators. Environmental Management, v. 33, p. 126–39, 2004.
FERREIRA, B. P.; MAIDA, M. Monitoramento dos Recifes de Coral no Brasil: Situação Atual e Perspectiva. 18. ed. Distrito Federal: Brasília, 2006.
FERREIRA, C. E. L.; GONÇALVES, J. E. A.; COUTINHO, R. Community structure of fishes and habitat complexity on a tropical rocky shore. Environmental Biology of Fishes, v. 61, p. 353–369, 2001.
FONSECA-KRUEL, V. S.; PEIXOTO, A. L. Etnobotânica na Reserva Extrativista Marinha de Arraial do Cabo, RJ, Brasil. Acta Botanica Brasilica, v. 18, n. 1, p. 177-190, 2004.
GLYNN, P. W. Coral reef bleaching: ecological perspective. Coral Reefs, v.129, n.1, p. 1-17, 1993.
GUIMARAENS, M. A.; COUTINHO, R. Spatial and temporal variation of benthic marine algae at the Cabo Frio upwelling region Rio de Janeiro, Brazil. Aquatic Botany, v. 52, p. 283-299, 1996.
GUZNER, B. et al. Indirect impacts of recreational scuba diving: Patterns of growth and predation in branching stony corals. Bulletin of Marine Science, v. 85, p. 727-42, 2010.
HISSA, D. C. et al. Efeito da temperatura no branqueamento de corais: Avaliação de Potências Bioindicadores do Aquecimento Global. Arquivos de Ciências do Mar, v. 42, n.2, p. 50-54, 2009.
HOEGH-GULDBERG, O. Climate change, coral bleaching and the future of the world’s coral reefs. Marine and Freshwater Research, v. 50, p. 839–866, 1999.
HUGHES, T. P. et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science, v. 359, p. 80–83, 2018.
HUGHES, T. P. et al. Global warming impairs stock-recruitment dynamics of corals. Nature, v.568, p. 387-390, 2019.
KARAKO-LAMPERT, S. et al. Do clades of symbiotic dinoflagellates in scleractinian corals of the Gulf of Eilat (Red Sea) differ from those of other coral reefs? Journal of Experimental Marine Biology and Ecology, v. 311, p. 301-314, 2004.
KELMO, F. Caracterização do branqueamento de corais no recife costeiro da praia de guarajuba, litoral norte do estado da Bahia. 1998. 104 f. Dissertação de Mestrado (Mestre em Ciência Geologia) - Universidade Federal da Bahia, Salvador, 1998.
KEMP, D. W. et al. Comparison of the thermal bleaching responses of the zoanthid Palythoa caribaeorum from three geographically different regions in south Florida. Journal of Experimental Marine Biology and Ecology, v. 335, p. 266 - 276, 2006.
LABOREL, J. Les peuplements de madréporaires des côtes tropicales du Brésil. Annales de l’Université d’Abidjan, sér. E, v. 2, n. 3, p. 1-260, 1970.
LANG, J. et al. Atlantic and Gulf rapid reef assessment, Protocols Version 5.4. 2010. Acesso em: 26 jan. 2019.
LEÃO, Z. M. A. N. et al. Status of Eastern Brazilian coral reefs in time of climate changes. Pan-American Journal of Aquatic Sciences, v. 5, n. 2, p. 52-63, 2010.
LEÃO, Z. M. A. N.; KIKUCHI, R. K. P.; TESTA, V. Corals and coral reefs of Brazil. In: Cortés, J. Latin American Coral Reefs. Amsterdam: Elsevier, 2003. Cap. 1.
LEÃO, Z. M. A. N.; KIKUCHI, R. K. P.; OLIVEIRA, M. D. M. Branqueamento de corais nos recifes da Bahia e sua relação com eventos de anomalias térmicas nas águas superficiais do oceano. Biota Neotropica, v. 8, n. 3, p. 69-82, 2008.
LESSER, M. P. et al. Bleaching in coral reef anthozoans: effects of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs, v. 8, p. 225-232, 1990.
LIMA, L. F. O.; COUTINHO, R. The reef coral Siderastrea stellata thriving at its range limit: population structure in Arraial do Cabo, southeastern Brazil. Bulletin of Marine Science, v. 92, n. 1, p. 107-121, 2016.
LONGO, G. O.; CORREIA, L. F. C.; MELLO, T. J. Coral recovery after a burial event: insights on coral resilience in a marginal reef. Marine Biodiversity, v. 50, p. 92, 2020.
LUNA-PÉREZ, B.; VALLE-PÉREZ, C.; SÁNCHEZ-LIZASO, J. L.Halocynthia papillosa as scuba diving impact indicator: An in situ experiment. Journal of Experimental Marine Biology and Ecology, v. 398, p. 33-39, 2011.
MACEDO, C. M. R. Microanatomia e Sistemática das espécies de Zoanthus Lamarck, 1801 (Cnidaria, Anthozoa, Zoanthidae) do litoral e ilhas oceânicas do Brasil. 1986. 141f. Dissertação de Mestrado (Mestre em Zoologia) - Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 1986.
MELO, L. V. et al. Ampliação do Porto do Forno na Reserva Extrativista Marinha em Arraial do Cabo, RJ. Boletim do Observatório Ambiental Alberto Ribeiro Lamego, v. 3, n. 2, p. 163-186, 2009.
MIES, M. et al. South Atlantic coral reefs are major global warming refugia and less susceptible to bleaching. Frontiers in Marine Science, v. 7, p. 1-13, 2020.
MIRANDA, R. J.; CRUZ, I. C. S.; LEÃO, Z. M. A. N. Coral bleaching in the Caramuanas reef (Todos os Santos Bay, Brazil) during the 2010 El Niño event. Latin American Journal of aquatic research, v. 41, n. 2, p. 351-360, 2013.
MOBERG, F.; FOLKE, C. Ecological goods and services of coral reef ecosystems. Ecological Economics, v. 29, p. 215-233, 1999.
OIGMAN-PSZCZOL, S. S.; FIGUEIREDO, M. A. O.; CREED, J. C. Distribution of benthic communities on the tropical rocky subtidal of Armação dos Búzios, Southeastern Brazil. Marine Ecology, v. 25, n. 3, p. 173-190, 2004.
OIGMAN-PSZCZOL, S. S.; CREED, J. C. Size structure and spatial distribution of corals Mussismilia hispida and Siderastrea stellata (Scleractinia) at Armação dos Búzios, Brazil. Bulletin of Marine Science, v. 74, n. 2, p. 433-448, 2004.
ORNELLAS, A. B.; COUTINHO, R. Spatial and temporal patterns of distribution and abundance of a tropical fish assemblage in a seasonal Sargassum bed, Cabo Frio Island, Brazil. Journal of Fish Biology, v. 53 Suppl A, p. 198-208, 1998.
PICCIANI, N. et al. Geographic patterns of Symbiodinium diversity associated with the coral Mussismilia hispida (Cnidaria, Scleractinia) correlate with major reef regions in the Southwestern Atlantic Ocean. Marine Biology, v. 163, p. 236, 2016.
POONIAN, C.; DAVIS, P. Z. R.; MCNAUGHTON, C. K. Impacts of recreational divers on palauan coral reefs and options management. Pacific Science, v. 4, p. 557-565, 2010.
RAMOS, C. A. C. et al. Quantification of reef benthos communities and variability inherent to the monitoring using video transect method. Environmental Monitoring and Assessment, v. 162, p. 95-101, 2010.
RODGERS, K. S. et al. Patterns of bleaching and mortality following widespread warming events in 2014 and 2015 at the Hanauma Bay Nature Preserve, Hawai’i. PeerJ, v. 5, p. 1-13, 2017.
ROGERS, R. et al. Coral health rapid assessment in marginal reef sites. Marine Biology Research, v. 10, n. 6, p. 612-624, 2014.
SANTOS, G. S. et al. Response of the zooxanthellae of Palythoa caribaeorum (Cnidaria: Zoanthidea) to different environmental conditions in coastal and oceanic ecosystems of the Tropical Atlantic. Helgoland Marine Research, v. 70, p. 2, 2016.
SASSI, C. F. C. et al. Pigmentation patterns of Siderastreastellata Verrill, 1868 (Cnidaria, Scleractinia) from coastal reefs in Northeastern Brazil and its relation with zooxanthellae and other microsymbionts. Pan-American Journal of Aquatic Sciences, v. 9, n. 3, p. 207-222, 2014.
SOUZA, C. A.; AMARAL, F. D. Variação morfométrica de algumas espécies de corais Mussidae (CNIDARIA, ANTHOZOA) do Brasil. Tropical Oceanography, v. 30, n. 2, p. 23-36, 2002.
SOUZA, I. M. M. Avaliação da cobertura e monitoramento do branqueamento de corais nos recifes de Maracajaú, RN. 2013. 61 f. Dissertação de Mestrado (Mestre em Ecologia) - Universidade Federal do Rio Grande do Norte, Natal, Rio Grande do Norte, 2013.
TEIXEIRA, C. D. et al. Sustained mass coral bleaching (2016–2017) in Brazilian turbid-zone reefs: taxonomic, cross-shelf and habitat-related trends. Coral Reefs, v. 38, p. 801–813, 2019.
VALENTIN, J. L. 1984. Analyse des paramètres hydrobiologiques dans la remontée de Cabo Frio (Brésil). Marine Biology, v. 82, p. 259-276, 1984.
ZAR. J. H. Biostatistical Analysis. New Jersey, USA: Prentice-Hall, 1999. 663 p.
Notas de autor
Información adicional
COMO CITAR (ABNT): SANTOS, L. A. et al. Branqueamento de corais e outros cnidários bentônicos no costão rochoso da Praia do Forno (Arraial do Cabo, RJ) durante as anomalias térmicas das águas superficiais do oceano ocorridas nos meses de fevereiro e maio de 2019. Vértices (Campos dos Goitacazes), v. 23, n. 2, p. 560-579, 2021. DOI: https://doi.org/10.19180/1809-2667.v23n22021p560-579. Disponível em: https://www.essentiaeditora.iff.edu.br/index.php/vertices/article/view/15892.
COMO CITAR (APA): Santos, L. A., Silva, B. C. A., Silva, K. C. R., Santos, R. C., Sousa, E. M., Muniz, R. A. & Barbosa, A. B. (2021). Branqueamento de corais e outros cnidários bentônicos no costão rochoso da Praia do Forno (Arraial do Cabo, RJ) durante as anomalias térmicas das águas superficiais do oceano ocorridas nos meses de fevereiro e maio de 2019. Vértices (Campos dos Goitacazes), 23(2), 560-579. https://doi.org/10.19180/1809-2667.v23n22021p560-579